
di PAOLO AJMONE MARSAN
 |
| Figura 1 - Tori di razza Blu Belga a sinistra e Piemontese a destra. |
Riassunto
La maggior parte delle mutazioni mendeliane identificate e caratterizzate nelle specie zootecniche sono causa di difetti genetici. Gli animali portatori di tali varianti sono di conseguenza esclusi dai programmi di selezione. Altre mutazioni, purtroppo poche, danno origine ad alleli in grado di migliorare alcuni caratteri di interesse. In questa breve rassegna ne vengono descritte tre, controllate da singoli geni che modificano caratteristiche interessanti per i bovini da latte e da carne: la muscolosità, che influenza l’efficienza della produzione di carne e la resa al macello, la presenza/assenza di corna, carattere associato al benessere animale e alla sicurezza degli operatori e la tolleranza al caldo, carattere sempre più interessante in un periodo di rapidi cambiamenti climatici. L’utilizzo di queste varianti nei programmi di miglioramento genetico è semplice se sono già presenti in alcuni individui all’interno della razza in selezione, più complesso nel caso in cui debbano essere introdotti da una razza diversa. In quest’ultimo caso le nuove tecnologie di gene-editing sarebbero di grande aiuto per inserire rapidamente le varianti favorevoli direttamente in animali di alto valore genetico.
Abstract
Genetics of mono-factorial traits and assisted selection in cattle Most of the Mendelian mutations identified and characterized in livestock species cause genetic defects. Animals carrying such variants are therefore excluded from breeding programmes. Other mutations, unfortunately few, give rise to alleles capable of improving some characters of interest. In this brief review, three are described, controlled by individual genes that modify interesting characteristics in dairy and beef cattle: double muscling, which influences the efficiency of meat production and slaughter yield, the presence/absence of horns, trait associated with animal welfare and operator safety, and tolerance to heat, an increasingly interesting character in a period of rapid climate change. The use of these variants in breeding programs is simple if they are already present in some individuals within the breed being selected, more complex in case they are to be introduced from a different breed. In the latter case, new gene-editing technologies would greatly speed-up the insertion of favorable variants directly in animals of high genetic value.
INTRODUZIONE
Sono trascorsi 200 anni dalla nascita di Mendel. Il fatto che i genetisti chiamino ancora mendeliani i caratteri controllati da singoli geni che segregano indipendentemente è indicativo dell’importanza della ricerca del monaco Agostiniano. Che i suoi risultati non siano stati riconosciuti e che le sue leggi siano state riscoperte da De Vries, Correns, Tschermak e von Seysenegg ben un ventennio più tardi è indicatore di quanto fosse avanzata la sua ricerca, che ha per la prima volta unito biologia e analisi dei dati raccolti, il primo esempio di interdisciplinarietà e importanza dell’analisi matematico-statistica in biologia. Nel settore zootecnico diversi caratteri sono controllati da singoli geni e segregano in modo mendeliano.
Il database OMIA (Online Mendelian Inheritance in Animals) ne elenca 292, due terzi dei quali controllati da varianti geniche note (Tabella 1). Una frazione di quanto noto nell’uomo e riportato nell’omologo database OMIM (Online Mendelian Inheritance in Man), che contiene più di 7000 fenotipi e 4000 varianti causative note. Per entrambe le specie più del 90% dei record si riferisce a varianti deleterie, che se presenti allo stato omozigote, o occasionalmente allo stato eterozigote, sono causa di alterazioni più o meno gravi. Nei bovini i fenotipi alterati riguardano principalmente la fertilità, il metabolismo, la morfologia ed il sistema immunitario.
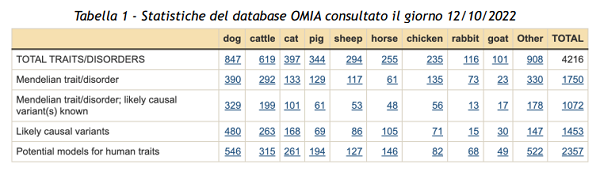
Sempre in questa specie, il rimanente 10% include varianti dei geni che codificano per le proteine del latte (caseine e sieroproteine), per il colore del mantello, per i gruppi sanguigni, per la muscolosità, la presenza/assenza di corna e per la termotolleranza. Mi soffermerò su questi ultimi tre aspetti perché di interesse per il miglioramento genetico in termini di aumento della sostenibilità delle produzioni e impatto sul benessere animale. Invito il lettore interessato a visitare il sito http://www.lgscr.it/ per esaminare tutti i test genetici offerti e spesso effettuati sulle razze bovine allevate nel nostro paese.
LA DOPPIA COSCIA
Torniamo al primo dei caratteri mendeliani da discutere, la muscolosità. Diverse varianti nel gene GDF-8, che codifica per la proteina miostatina hanno come effetto l’aumento della muscolosità degli animali, in particolare nella parte posteriore della groppa. Questo effetto, inizialmente scoperto nel topo (McFerron et al., 1997), si è poi rivelato il responsabile di un fenotipo analogo nei bovini (Marin et al., 1997). Il fenotipo causato dalla mutazione è noto come “doppia coscia”, “doppia groppa” o “ipertrofia muscolare”. In realtà l’incremento della muscolosità è causato da un’iperplasia dei muscoli, cioè ad un aumentato numero di cellule muscolari. Oggi sappiamo che la mutazione ha influenza anche su metabolismo glucidico e lipidico, sulla produzione di bile (Wu et al., 2022) e, almeno nel topo, sulla funzionalità dei mitocondri (Gu et al., 2022). L’iperplasia è causata dal malfunzionamento della miostatina, appunto codificata dal gene GDF-8, che nel bovino mappa sul cromosoma 2 (BTA2).
Questa proteina si lega ad un recettore in forma di dimero, e porta alla formazione di un complesso che interagisce con il DNA e attiva una serie di geni che in ultima analisi reprimono la divisione delle cellule muscolari durante l’ultima fase della gravidanza. Il dominio fondamentale per la dimerizzazione è ricco di cisteine responsabili dell’interazione dei due monomeri di proteina che si legano al recettore (Bongiorni et al., 2016). Diverse mutazioni che troncano la proteina in questo dominio sono state catturate dall’uomo nel corso della storia evolutiva dei bovini europei e sono oggi presenti in diverse razze da carne. Il fenotipo risultante è tanto più estremo quanto più la mutazione riduce il numero di cisteine, e quindi altera la capacità della proteina di formare dimeri.
Le mutazioni mostrano una dominanza parziale, gli eterozigoti mostrano un fenotipo iperplasico ma non al livello degli omozigoti. È ipotizzabile ciò sia dovuto al fatto che la proteina agisce come dimero. Animali eterozigoti producono sia la forma corretta che quella alterata di miostatina e solo un quarto dei possibili dimeri, quello formato da due forme corrette, funziona normalmente, mentre gli altri hanno efficacia ridotta. La razza che sicuramente esprime il fenotipo in modo più estremo è la Blu Belga (Fig. 1), spesso usata per fecondare bovine da latte non destinate alla rimonta o in incrocio con altre razze da carne. In questa razza la mutazione è fissata, cioè tutti gli animali sono omozigoti per la mutazione. Il fenotipo estremo mette a rischio le bovine di Blu Belga durante il parto che quasi sempre è cesareo. Per questo motivo in condizioni naturali questa mutazione sarebbe scomparsa velocemente a causa degli effetti negativi sulla fitness dei portatori.
La mutazione è anche presente in due razze italiane, nella Piemontese, nota appunto come Piemontese della Coscia, e nella Marchigiana. I riproduttori maschi Piemontesi hanno la mutazione allo stato omozigote, mentre le femmine sono omozigoti per l’allele selvatico, cioè non mutato, e sono selezionate per la facilità di parto. Ciò, insieme al fatto che i vitelli sono eterozigoti per la mutazione e che la mutazione posseduta dalla Piemontese non cancella completamente la serie di cisteine, limita l’aumento di muscolosità alla parte posteriore degli animali e riduce moltissimo la necessità di parti cesarei nella razza.
Nella Marchigiana la mutazione è stata scoperta grazie a una collaborazione tra l’associazione allevatori delle razze bianche italiane (ANABIC) e l’Università della Tuscia (Marchitelli et al., 2003). La mutazione Marchigiana causa un fenotipo ancor meno estremo di quello della Piemontese ed è ancora poco diffusa nella razza. L’associazione deve infatti contemporaneamente aumentarne la frequenza ed evitare un’eccesiva consanguineità, quest’ultima associata ad anomalie genetiche ed ipofertilità. Gli animali “ipertrofici” hanno caratteristiche favorevoli, carne più magra, maggior resa al macello e trasformano in modo più efficiente gli alimenti in muscolo. Sono quindi più efficienti, una caratteristica che aumenta la sostenibilità del loro allevamento. D’altra parte, l’efficienza contrasta con la necessità di cure veterinarie e con il loro benessere che nei casi più estremi, come quello della Blu Belga, può dare adito a parti distocici.
L’ASSENZA DI CORNA
Il secondo carattere mendeliano di interesse per la selezione è l’assenza/presenza di corna. L’assenza di corna è un carattere desiderabile, sia per la sicurezza degli operatori, sia degli stessi animali, che durante l’instaurarsi delle relazioni gerarchiche, tipiche delle specie che vivono in gruppo, possono ferirsi. Inoltre, l’assenza “genetica” delle corna eviterebbe la pratica della decornazione dei vitelli, con vantaggi per il benessere animale. Il fenotipo assenza è dominante sulla presenza, cioè animali eterozigoti per la mutazione sono acorni come quelli omozigoti, mentre gli animali con le corna sono omozigoti per l’allele selvatico (Fig. 2). La mutazione è definita “polled”, dall’inglese ed è presente in diverse razze, tra cui: Holstein, Jersey, Limousine, Charolaise, Blond d’Aquitaine e Aberdeen Angus. Quest’ultima è fissata e selezionata per la mutazione acorne, i riproduttori non vengono abilitati alla monta se presentano corna. Mutazioni in loci diversi provocano l’assenza delle corna, alcune di queste provocano o sono associate a difetti genetici, per cui non possono essere selezionate a favore ed introdotte nei programmi di breeding (Gehrke et al., 2020). Le mutazioni presenti sul cromosoma 1 non hanno effetti secondari indesiderati (Stuckey et al., 1996) e possono essere sfruttate nei programmi di selezione.
 |
| Figura 2 - Tori di razza Hereford con corna a sinistra e senza corna perché portatori della mutazione “polled Celtica” sul cromosoma 1. |
DNA (Allais-Bonet et al., 1013), PF è determinato da una duplicazione in tandem di 80 nucleotidi (Rothammer et al., 2014). Le mutazioni non sono in porzioni del genoma dove risiedono geni codificanti. È stato ipotizzato che influenzino l’espressione di RNA non codificanti importanti per la formazione dei primi abbozzi delle corna durante lo sviluppo embrionale (Simon et al., 2022), in realtà i meccanismi che collegano le mutazioni al fenotipo acorne non sono ancora stati ben compresi. Anche in questo caso nelle razze in cui la mutazione è rara, è necessario che i programmi di accoppiamento per aumentarne la frequenza siano ben progettati, per evitare un eccesso di consanguineità e mantenere la biodiversità necessaria per il miglioramento di altri caratteri oggetto di selezione.
LA MUTAZIONE “SLICK”
Il terzo fenotipo mendeliano che vorrei discutere è apparentemente poco importante ma sostanzialmente interessantissimo per la futura sostenibilità, oserei dire, dell’intero settore zootecnico. Il fenotipo è denominato “slick”, letteralmente “liscio”, per la presenza di un mantello a pelo cortissimo e appunto liscio (Fig. 3).
 |
| Figura 3. Confronto tra un bovino portatore della mutazione slick (a sinistra) e un bovino omozigote per l’allele selvatico (a destra). |
La mutazione è dominante e induce uno stop prematuro nella coda citoplasmatica del gene PRLR che codifica per il recettore dell’ormone prolattina (Littlejohn et al., 2014). Oltre al mantello “liscio” la mutazione sembra sia responsabile di altri cambiamenti fisiologici, come una maggiore capacità di sudorazione (Dikmen et al., 2014), che rendono gli animali “lisci” più tolleranti alle alte temperature rispetto a quelli omozigoti per l’allele selvatico. La mutazione è stata inizialmente scoperta in due razze sudamericane, di origine spagnola, Senepol e Carora (Olson et al., 2003), un esempio paradigmatico dell’utilità di conservare razze locali adatte ad ambienti difficili, anche se non molto produttive. Utilizzando incroci ricorrenti, la mutazione è stata trasferita con successo nella razza Holstein, cioè la Frisona americana, cosmopolita, e ha dimostrato di indurre termotolleranza anche in questo background genetico. Durante l’estate della Florida, calda e umida, le bovine Holstein in lattazione portatrici della mutazione hanno mantenuto una temperatura vaginale inferiore di 0,6°C rispetto alle bovine non mutate (Carmickle et al., 2022).
Questo dato è interessante perché il caldo estivo è causa di stress per le bovine ad alta produzione e determina una netta perdita di produttività a causa di una minore ingestione (Bernabucci et al., 2014). La perdita è evidente soprattutto quando lo stress da caldo coincide con il periodo del periparto, periodo durante il quale la produzione di latte cresce rapidamente e non è accompagnata da un analogo aumento di capacità di ingestione. In conseguenza di questo fatto le bovine vanno incontro ad un bilancio energetico negativo e devono mobilizzare le riserve corporee per far fronte alle richieste energetiche necessarie per mantenere la produzione di latte. Al contrario delle bovine “wild type”, le bovine “slick” hanno mostrato la stessa capacità produttiva indipendentemente dalla stagione di parto (inverno o estate; Dikmen et al., 2014). Il mantenimento della produzione estiva è stato osservato sia in ambienti caldo-umidi, quali quelli della Florida, che caldo-secchi, come in California (Carmickle et al., 2022). Recentemente sono state scoperte nuove varianti nel gene del recettore della prolattina in altre razze, sempre dei Caraibi (Flores et al., 2021). Tutte inducono codoni di stop prematuro e di conseguenza proteine tronche, e tutte sono associate a tolleranza a stress da alte temperature.
UTILIZZO DI CARATTERI MENDELIANI NEI PROGRAMMI DI SELEZIONE
Come fare ad integrare questi geni nei programmi di selezione? Abbiamo a disposizione fondamentalmente due strategie: la prima è quella dell’uso su larga scala di riproduttori portatori delle mutazioni nell’ambito di un programma di miglioramento genetico, mentre la seconda è il ricorso all’editing del genoma. Nel primo caso è necessario che le associazioni di razza interessate definiscano un programma combinato di miglioramento all’interno del quale inserire tori che possiedano i geni di interesse, aumentandone la frequenza, mantenendo un buon livello genetico dei portatori e facendo attenzione a conservare la biodiversità della razza. Nel secondo si tratterebbe, invece, di effettuare una modificazione genetica inserendo la mutazione nel genoma degli animali attraverso “gene editing” (Doudna e Charpentier, 2014). In questo caso la mutazione potrebbe essere inserita in modo “chirurgico” anche in razze che non la possiedono. Un confronto tra i due approcci ha mostrato come velocità e costi siano minori con l’applicazione del gene editing, rispetto all’approccio tradizionale (Mueller et al., 2021). Tutte le mutazioni prima descritte sono state oggetto di gene editing. Diverse specie sono state “editate” al fine di disattivare il gene della miostatina: bovini (Proudfoot et al., 2015; Luo et al., 2017; Gim et al., 2022a) ovini Proudfoot et al., 2015), capre (Kalds et al., 2022) suini (Proudfoot et al., 2015; Wang et al., 2015) e conigli (Zheng et al., 2022). Gli interventi di editing hanno ottenuto i risultati desiderati e nei bovini è stata dimostrata la trasmissibilità dell’intervento di editing della miostatina alla generazione successiva. (Gim et al., 2022b). L’editing della mutazione polled ha invece avuto esiti diversi, almeno fino ad ora. Alcuni interventi hanno avuto successo, dimostrando la trasmissione della mutazione introdotta e del fenotipo acorne (Young et a., 2021), altri hanno introdotto il fenotipo, ma i nati non sono sopravvissuti (Schuster et al., 2021), altri ancora hanno confermato la presenza della mutazione introdotta ma non hanno dato origine al fenotipo (Hennig et al., 2022). La complessità delle varianti esistenti e la mancanza di conoscenza dei complessi meccanismi molecolari che determinano l’insorgenza e lo sviluppo degli abbozzi delle corna a livello embrionale non hanno per ora permesso di ottenere risultati consistenti. Anche la mutazione slick è stata oggetto di editing. È stato prodotto un modello murino (Liu et al., 2022) che si pensa permetterà di studiare in maggior dettaglio i meccanismi fisiologici che determinano la tolleranza al caldo osservata nei bovini, sebbene dal punto di vista biologico le due specie siano profondamente diverse. Nei bovini l’editing del gene PRLR è in corso, anche se i risultati non sono ancora pubblicati (Tad Sonstegard – Accelligen-Recombinetics, comunicazione personale 2022).
L’intervento è previsto nell’ambito di un progetto dedicato al miglioramento della zootecnica africana finanziato dalla fondazione Bill e Melinda Gates. Nello stesso progetto sono programmati interventi di editing anche per migliorare la produzione di latte e la resistenza alla tubercolosi. Nell’Unione Europea questa strada non risulta ancora perseguibile, mentre sarebbe una via da adottare per poter inserire varianti favorevoli con rapidità e direttamente in animali di alto valore genetico. Da questo punto di vista varianti associate all’adattamento ai cambiamenti climatici, come la mutazione slick, e alla resistenza alle malattie sarebbero prioritarie. I rischi associati a questa tecnologia sono minimi considerato che le modificazioni sono di una o pochi nucleotidi e che sono già avvenute durante la storia evolutiva della specie. Inoltre, i risultati dell’editing sono facilmente controllabili attraverso sequenziamento, che permette di identificare la presenza di eventuali variazioni “off target” e stimarne gli effetti (Zhou et al., 2019). Mentre l’Europa riflette, diversi altri Paesi hanno adottato la tecnologia e la stanno usando per accelerare il miglioramento genetico in diverse specie, animali e vegetali.
REFERENZE
Allais-Bonnet A., Grohs C., Medugorac I., Krebs S., Djari A., Graf A., Fritz S., Seichter D., Baur A., Russ I., et al. Novel insights into the bovine polled phenotype and horn ontogenesis in Bovidae. PLoS ONE. 2013;8:e63512.
Bernabucci U, Biffani S, Buggiotti L, Vitali A, Lacetera N, Nardone A. The effects of heat stress in Italian Holstein dairy cattle. J Dairy Sci. 2014;97(1):471-86. doi: 10.3168/jds.2013-6611. Epub 2013 Nov 7. PMID: 24210494.
Bongiorni S, Valentini A, Chillemi G. Structural and Dynamic Characterization of the C313Y Mutation in Myostatin Dimeric Protein, Responsible for the "Double Muscle" Phenotype in Piedmontese Cattle. Front Genet. 2016 Feb 11;7:14. doi: 10.3389/fgene.2016.00014. PMID: 26904102; PMCID: PMC4749705.
Carmickle AT, Larson CC, Hernandez FS, Pereira JMV, Ferreira FC, Haimon MLJ, Jensen LM, Hansen PJ, Denicol AC. Physiological responses of Holstein calves and heifers carrying the SLICK1 allele to heat stress in California and Florida dairy farms. J Dairy Sci. 2022 Nov;105(11):9216-9225. doi: 10.3168/jds.2022-22177. Epub 2022 Sep 14. PMID: 36114060.
Dikmen S, Khan FA, Huson HJ, Sonstegard TS, Moss JI, Dahl GE, Hansen PJ. The SLICK hair locus derived from Senepol cattle confers thermotolerance to intensively managed lactating Holstein cows. J Dairy Sci. 2014 Sep;97(9):5508-20. doi: 10.3168/jds.2014-8087. Epub 2014 Jul 2. PMID: 24996281.
Dikmen, S., E. Alava, E. Pontes, J. M. Fear, B. Y. Dikmen, T. A. Olson, and P. J. Hansen. 2008. Differences in thermoregulatory ability between slick-haired and wild-type lactating Holstein cows in response to acute heat stress. J. Dairy Sci. 91:3395–3402. https: //doi.org/10.3168/jds.2008-1072. 122
Doudna JA, Charpentier E. Genome editing. The new frontier of genome engineering with CRISPRCas9. Science. 2014 Nov 28;346(6213):1258096. doi: 10.1126/science.1258096. PMID: 25430774.
Flórez Murillo, J. M., A. J. Landaeta-Hernández, E. S. Kim, J. R. Bostrom, S. A. Larson, A. M. Pérez O’Brien, M. A. Montero-Urdaneta, J. F. Garcia, and T. S. Sonstegard. 2021.
Three novel nonsense mutations of prolactin receptor found in heat-tolerant Bos taurus breeds of the Caribbean Basin. Anim. Genet. 52:132–134. https://doi.org/10.1111/age.13027.
Gehrke, L.J., Upadhyay, M., Heidrich, K. et al. A de novo frameshift mutation in ZEB2 causes polledness, abnormal skull shape, small body stature and subfertility in Fleckvieh cattle. Sci Rep 10, 17032 (2020). https://doi.org/10.1038/s41598-020-73807-5.
Gim GM, Kwon DH, Eom KH, Moon J, Park JH, Lee WW, Jung DJ, Kim DH, Yi JK, Ha JJ, Lim KY, Kim JS, Jang G. Production of MSTN-mutated cattle without exogenous gene integration using CRISPR-Cas9. Biotechnol J. 2022b Jul;17(7):e2100198. doi: 10.1002/biot.202100198. Epub 2021 Jul 22. PMID: 34247443.
Gim GM, Uhm KH, Kwon DH, Kim MJ, Jung DJ, Kim DH, Yi JK, Ha JJ, Yum SY, Son WJ, Lee JH, Park JH, Song KY, Lee WW, Jang G. Germline transmission of MSTN knockout cattle via CRISPR-Cas9. Theriogenology. 2022a Oct 15;192:22-27. doi: 10.1016/j.theriogenology.2022.08.021. Epub 2022 Aug 18. PMID: 36037573.
Grobet L., Martin L.J., Poncelet D., Pirottin D., Brouwers B., Riquet J., Schoeberlein A., Dunner S., Ménissier F., Massabanda J., Fries R., Hanset R., Georges M. A deletion in the bovine myostatin gene causes the double-muscled phenotype in cattle. Nat Genet. 1997 Sep;17(1):71- 4. doi: 10.1038/ng0997-71. PMID: 9288100.
Gu M, Wei Z, Wang X, Gao Y, Wang D, Liu X, Bai C, Su G, Yang L, Li G. Myostatin Knockout Affects Mitochondrial Function by Inhibiting the AMPK/SIRT1/PGC1α Pathway in Skeletal Muscle. Int J Mol Sci. 2022 Nov 8;23(22):13703. doi: 10.3390/ijms232213703. PMID: 36430183.
Hennig, S.L., Owen, J.R., Lin, J.C. et al. A deletion at the polled PC locus alone is not sufficient to cause a polled phenotype in cattle. Sci Rep 12, 2067 (2022). https://doi.org/10.1038/s41598-022-06118-6.
K. Wang, H. Ouyang, Z. Xie, C. Yao, N. Guo, M. Li, Jiao, H., Pang, D. Efficient generation of myostatin mutations in pigs using the CRISPR/Cas9 system Sci Rep, 5 (2015), Article 16623.
Kalds P, Crispo M, Li C, Tesson L, Anegón I, Chen Y, Wang X, Menchaca A. Generation of DoubleMuscled Sheep and Goats by CRISPR /Cas9-Mediated Knockout of the Myostatin Gene. Methods Mol Biol. 2022;2495:295-323. doi: 10.1007/978-1-0716-2301-5_16. PMID: 35696040.
Littlejohn, M. D., K. M. Henty, K. Tiplady, T. Johnson, C. Harland, T. Lopdell, R. G. Sherlock, W. Li, S. D. Lukefahr, B. C. Shanks, D. J. Garrick, R. G. Snell, R. J. Spelman, and S. R. Davis. 2014. Functionally reciprocal mutations of the prolactin signalling path way define hairy and slick cattle. Nat. Commun. 5:5861. https://doi.org/10.1038/ncomms6861.
Liu J, Verma PJ. Generating a Heat-Tolerance Mouse Model. Methods Mol Biol. 2022;2495:259-272. doi: 10.1007/978-1-0716-2301-5_14. PMID: 35696038. Luo J, Song Z, Yu S, Cui D, Wang B, et al. (2014) Efficient Generation of Myostatin (MSTN) Biallelic Mutations in Cattle Using Zinc Finger Nucleases. PLOS ONE 9(4): e95225. https://doi.org/10.1371/journal.pone.0095225.
Marchitelli C, Savarese MC, Crisà A, Nardone A, Ajmone Marsan P, Valentini A. Double muscling in Marchigiana beef breed is caused by a stop codon in the third exon of myostatin gene. Mamm Genome. 2003 Jun;14(6):392-5. doi: 10.1007/s00335-002-2176-5. PMID: 12879361.
McPherron, A.C.; Lawler, A.M.; Lee, S.J. Regulation of skeletal muscle mass in mice by a new TGFbeta superfamily member. Nature 1997, 387, 83–90.
Mueller ML, Cole JB, Connors NK, Johnston DJ, Randhawa IAS, Van Eenennaam AL. Comparison of Gene Editing Versus Conventional Breeding to Introgress the POLLED Allele Into the Tropically Adapted Australian Beef Cattle Population. Front Genet. 2021 Feb 11;12:593154. doi: 10.3389/fgene.2021.593154. PMID: 33643378; PMCID: PMC7905321. 123
Olson TA, Lucena C, Chase CC Jr, Hammond AC. Evidence of a major gene influencing hair length and heat tolerance in Bos taurus cattle. J Anim Sci. 2003 Jan;81(1):80-90. doi: 10.2527/2003.81180x. PMID: 12597376.
Proudfoot, C., Carlson, D.F., Huddart, R. et al. Genome edited sheep and cattle. Transgenic Res 24, 147–153 (2015). https://doi.org/10.1007/s11248-014-9832-x.
Rothammer S., Capitan A., Mullaart E., Seichter D., Russ I., Medugorac I. The 80-kb DNA duplication on BTA1 is the only remaining candidate mutation for the polled phenotype of Friesian origin. Genet. Sel. Evol. 2014;46:44. doi: 10.1186/1297-9686-46-44.
Schuster, F., Aldag, P., Frenzel, A. et al. CRISPR/Cas12a mediated knock-in of the Polled Celtic variant to produce a polled genotype in dairy cattle. Sci Rep 10, 13570 (2020). https://doi.org/10.1038/s41598-020-70531-y.
Simon R, Drögemüller C, Lühken G. The Complex and Diverse Genetic Architecture of the Absence of Horns (Polledness) in Domestic Ruminants, including Goats and Sheep. Genes (Basel). 2022 May 6;13(5):832. doi: 10.3390/genes13050832. PMID: 35627216; PMCID: PMC9140736.
Stookey J.M., Goonewardene L.A. A comparison of production traits and welfare implications between horned and polled beef bulls. Can. J. Anim. Sci. 1996;76:1–5. doi: 10.4141/cjas96- 001.
Wu D, Gu M, Wei Z, Bai C, Su G, Liu X, Zhao Y, Yang L, Li G. Myostatin Knockout Regulates Bile Acid Metabolism by Promoting Bile Acid Synthesis in Cattle. Animals (Basel). 2022 Jan 15;12(2):205. doi: 10.3390/ani12020205. PMID: 35049827; PMCID: PMC8772948.
Young, A.E., Mansour, T.A., McNabb, B.R. et al. Genomic and phenotypic analyses of six offspring of a genome-edited hornless bull. Nat Biotechnol 38, 225–232 (2020). https://doi.org/10.1038/s41587-019-0266-0.
Zheng Y, Zhang Y, Wu L, Riaz H, Li Z, Shi D, Rehman SU, Liu Q, Cui K. Generation of Heritable Prominent Double Muscle Buttock Rabbits via Novel Site Editing of Myostatin Gene Using CRISPR/Cas9 System. Front Vet Sci. 2022 May 20;9:842074. doi: 10.3389/fvets.2022.842074. PMID: 35669173; PMCID: PMC9165342.h Zhou S, Cai B, He C, Wang Y, Ding Q, Liu J, Liu Y, Ding Y, Zhao X, Li G, Li C, Yu H, Kou Q, Niu W, Petersen B, Sonstegard T, Ma B, Chen Y, Wang X. Programmable Base Editing of the Sheep Genome Revealed No Genome-Wide Off-Target Mutations. Front Genet. 2019 Mar 15;10:215. doi: 10.3389/fgene.2019.00215. PMID: 30930940; PMCID: PMC6428697.
Figura1-(Fonti:www.chiacchierini.it/tori-da-carne/toro-da-carne-razza-bianca-blu-belga/www.sigilloitaliano.it/i-prodotti/il-fassone-di-razza-piemontese/la-razza-piemontese/
Figura2-Fonti:www.afs.ca.uky.edu/livestock/beef/breeds/Hereford, www.britannica.com/animal/Herefordbreed-of-cattl
Figura3-Fonte:www.researchgate.net/figure/Slick-coat-type-Photographs-contrasting-slick-and-nonslickSenepol-crossbreeds-The_fig4_269698991
Articolo tratto dagli atti del convegno organizzato dal MULSA
"Gregor Mendel il mendelismo e la genetica agraria ".

E' Professore ordinario di Miglioramento generico animale, Università Cattolica del Sacro Cuore, Piacenza.
Nessun commento:
Posta un commento